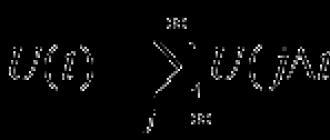Политенные хромосомы. Хромосомы в мейозе Конъюгация гомологичных хромосом
В два раза. Происходит в два этапа (редукционный и эквационный этапы мейоза). Мейоз происходит в половых клетках и связан с образованием гамет.
С уменьшением числа хромосом в результате мейоза в жизненном цикле происходит переход от диплоидной фазы к гаплоидной. Восстановление плоидности (переход от гаплоидной фазы к диплоидной) происходит в результате полового процесса .
В связи с тем, что в профазе первого, редукционного, этапа происходит попарное слияние (конъюгация) гомологичных хромосом , правильное протекание мейоза возможно только в диплоидных клетках или в чётных полиплоидах (тетра-, гексаплоидных и т. п. клетках). Мейоз может происходить и в нечётных полиплоидах (три-, пентаплоидных и т. п. клетках), но в них, из-за невозможности обеспечить попарное слияние хромосом в профазе I, расхождение хромосом происходит с нарушениями, которые ставят под угрозу жизнеспособность клетки или развивающегося из неё многоклеточного гаплоидного организма.
Этот же механизм лежит в основе стерильности межвидовых гибридов . Поскольку у межвидовых гибридов в ядре клеток сочетаются хромосомы родителей, относящихся к различным видам, хромосомы обычно не могут вступить в конъюгацию. Это приводит к нарушениям в расхождении хромосом при мейозе и, в конечном счете, к нежизнеспособности половых клеток, или гамет (основным средством борьбы с этой проблемой является применение полиплоидных хромосомных наборов, поскольку в данном случае каждая хромосома конъюгирует с соответствующей хромосомой своего набора). Определённые ограничения на конъюгацию хромосом накладывают и хромосомные перестройки (масштабные делеции , дупликации , инверсии или транслокации).
Фазы мейоза
Мейоз состоит из 2 последовательных делений с коротким периодом (интеркинез) между ними.
- Профаза I - профаза первого деления очень сложная и состоит из 5 стадий:
- Лептотена , или лептонема - упаковка хромосом, конденсация ДНК с образованием хромосом в виде тонких нитей (хромосомы укорачиваются).
- Зиготена , или зигонема - происходит конъюгация - соединение гомологичных хромосом с образованием структур, состоящих из двух соединённых хромосом, называемых тетрадами или бивалентами и их дальнейшая компактизация.
- Пахитена , или пахинема - (самая длительная стадия) - в некоторых местах гомологичные хромосомы плотно соединяются, образуя хиазмы . В них происходит кроссинговер - обмен участками между гомологичными хромосомами.
- Диплотена , или диплонема - происходит частичная деконденсация хромосом, при этом часть генома может работать, происходят процессы транскрипции (образование РНК), трансляции (синтез белка); гомологичные хромосомы остаются соединёнными между собой. У некоторых животных в ооцитах хромосомы на этой стадии профазы мейоза приобретают характерную форму хромосом типа ламповых щёток .
- Диакинез - ДНК снова максимально конденсируется, синтетические процессы прекращаются, растворяется ядерная оболочка; центриоли расходятся к полюсам; гомологичные хромосомы остаются соединёнными между собой.
К концу профазы I центриоли мигрируют к полюсам клетки, формируются нити веретена деления , разрушаются ядерная мембрана и ядрышки . Генетический материал - 2n4c (n - число хромосом, c - число молекул ДНК).
- Метафаза I - бивалентные хромосомы выстраиваются вдоль экватора клетки. Генетический материал - 2n4c.
- Анафаза I - микротрубочки сокращаются, биваленты делятся, и хромосомы расходятся к полюсам. Важно отметить, что, из-за конъюгации хромосом в зиготене, к полюсам расходятся целые хромосомы, состоящие из двух хроматид каждая, а не отдельные хроматиды, как в митозе . У каждого полюса генетический материал n2c, во всей клетке 2n4c.
- Телофаза I
В результате первого редукционного деления мейоза I образуется две клетки с генетическим материалом n2c
Второе деление мейоза следует непосредственно за первым, без выраженной интерфазы: S-период отсутствует, поскольку перед вторым делением не происходит репликации ДНК.
- Профаза II - происходит конденсация хромосом, клеточный центр делится и продукты его деления расходятся к полюсам ядра, разрушается ядерная оболочка, образуется веретено деления, перпендикулярное первому веретену.
- Метафаза II - унивалентные хромосомы (состоящие из двух хроматид каждая) располагаются на «экваторе» (на равном расстоянии от «полюсов» ядра) в одной плоскости, образуя так называемую метафазную пластинку .
- Анафаза II - униваленты делятся и хроматиды расходятся к полюсам.
- Телофаза II - хромосомы деспирализуются и появляется ядерная оболочка.
В результате из одной диплоидной клетки образуется четыре гаплоидных клетки с генетическим материалом nc. В тех случаях, когда мейоз сопряжён с гаметогенезом (например, у многоклеточных животных), при развитии яйцеклеток первое и второе деления мейоза резко неравномерны. В результате формируется одна гаплоидная яйцеклетка и три так называемых редукционных тельца (абортивные дериваты первого и второго делений).
Варианты
У некоторых простейших мейоз протекает иначе, чем описанный выше типичный мейоз многоклеточных . Например, может протекать только одно, а не два последовательных, мейотических деления, а кроссинговер - проходить во время другой фазы мейоза
КОНЪЮГАЦИЯ - Гаплоидные гаметы, образовавшиеся при делении диплоидной клетки путем мейоза, содержат по одной хромосоме каждой гомологичной пары (отцовского или материнского происхождения), т.е. только половину исходного числа хромосом. В связи с этим к аппарату клеточного деления здесь предъявляется дополнительное требование: гомологи должны "узнавать" друг друга и соединяться в пары, перед тем как они выстроятся на экваторе веретена. Такое спаривание, или конъюгация, гомологичных хромосом материнского и отцовского происхождения происходит только в мейозе. Во время первого деления мейоза происходит репликация ДНК, и каждая хромосома состоит после этого из двух хроматид, гомологичные хромосомы конъюгируют по всей своей длине, и между хроматидами спаренных хромосом происходит кроссинговер
КРОССИНГОВЕР (crossing-over): обмен генетического материала между хромосомами, как результат "разрыва" и соединения хромосом; процесс обмена участками хромосом при перекресте хромосом (рис. 118 , Б4).
Во время пахитены (стадия толстых нитей), гомологичные хромосомы находятся в состоянии конъюгации длительный период: у дрозофилы - четверо суток, у человека больше двух недель. Все это время отдельные участки хромосом находятся в очень тесном соприкосновении. Если в таком участке произойдет разрыв цепочек ДНК одновременно в двух хроматидах, принадлежащих разным гомологам, то при восстановлении разрыва может получиться так, что ДНК одного гомолога окажется соединенной с ДНК другой, гомологичной хромосомы. Этот процесс носит -название кроссинговера (англ. crossing-over - перекрест).
Поскольку кроссинговер - взаимный обмен гомологичными участками хромосом между гомологичными (парными) хромосомами исходных гаплоидных наборов - особи имеют новые, различающиеся между собой генотипы. При этом достигается перекомбинация наследственных свойств родителей, что увеличивает изменчивость и дает более богатый материал для естественного отбора.
Гены перемешиваются благодаря слиянию гамет двух различных особей, однако генетические изменения осуществляются не только этим путем. Никакие два потомка одних и тех же родителей (если только это не идентичные близнецы) не будут абсолютно одинаковыми. Во время мейоза осуществляются два различных вида пересортировки генов.
Один вид пересортировки - это результат случайного распределения разных материнских и отцовских гомологов между дочерними клетками при первом делении мейоза, каждая гамета получает свою, отличную от других выборку материнских и отцовских хромосом. Из этого следует, что клетки любой особи могут в принципе образовать 2 в степени n генетически различающихся гамет, где n - гаплоидное число хромосом. Однако на самом деле число возможных гамет неизмеримо больше из-за кросинговера (перекреста) - процесса, происходящего во время длительной профазы первого деления мейоза, когда гомологичные хромосомы обмениваются участками. У человека в каждой паре гомологичных хромосом кроссинговер происходит в среднем в 2 - 3 точках.
При кроссинговере происходит разрыв двойной спирали ДНК в одной материнской и одной отцовской хроматиде, а затем получившиеся отрезки воссоединяются "наперекрест" (процесс генетической рекомбинации). Рекомбинация происходит в профазе первого деления мейоза, когда две сестринские хроматиды так тесно сближены друг с другом, что их невозможно увидеть в отдельности. Гораздо позже в этой растянутой профазе становятся ясно различимы две отдельные хроматиды каждой хромосомы. В это время видно, что они связаны своими центромерами и тесно сближены по всей длине. Два гомолога остаются связанными в тех точках, где произошел кроссинговер между отцовской и материнской хроматидами. В каждой такой точке, которую называют хиазмой, две из четырех хроматид перекрещиваются Таким образом, это морфологический результат произошедшего кроссинговера, который сам по себе недоступен для наблюдения.
На этой стадии мейоза гомологи в каждой паре (или бивалент) остаются связанными друг с другом по меньшей мере одной хиазмой. Во многих бивалентах бывает большее число хиазм, так как возможны множественные перекресты между гомологами

Во-вторых, боковой конъюгации хроматид . Клетки, в которых есть политенные хромосомы, теряют способность к делению, они являются дифференцированными и активно секретирующими, то есть, политенизация хромосом является способом увеличения числа копий генов для синтеза какого-либо продукта. Политенные хромосомы можно наблюдать у двукрылых , у растений в клетках, связанных с развитием зародыша, у инфузорий при формировании макронуклеуса . Политенные хромосомы значительно увеличиваются в размерах, что облегчает их наблюдение и что позволяло изучать активность генов ещё в 1930-е годы. Принципиальным отличием от других типов хромосом является то, что политенные хромосомы являются интерфазными, тогда как все остальные можно наблюдать только во время митотического или мейотического деления клетки .
Классическим примером являются гигантские хромосомы в клетках слюнных желёз личинок Drosophila melanogaster . Репликация ДНК в этих клетках не сопровождается делением клетки , что приводит к накоплению вновь построенных нитей ДНК . Эти нити плотно соединены между собой по длине. Кроме того, в слюнных железах происходит соматический синапсис гомологичных хромосом, то есть, не только сестринские хроматиды конъюгируют между собой, но и гомологичные хромосомы каждой пары конъюгируют между собой. Таким образом, в клетках слюнных желёз можно наблюдать гаплоидное число хромосом .
История
Термин «политенная хромосома» предложил П. Коллер (P. Koller ) в 1935 году, а окончательно ввёл в науку С. Дарлингтон в 1937 году .
Размеры
Политенные хромосомы во много раз превышают по размеру хромосомы обычных соматических клеток . Они, как правило, в 100-200 раз длиннее и в 1000 раз толще (содержат до 1000 хромосом), чем хромосомы многих интерфазных клеток (как половых , так и соматических). Так, у личинок D. melangaster общая длина четырёх пар хромосом в слюнных железах составляет 2000 мкм, а в обычных соматических клетках эта величина равна 7,5 мкм .
Строение
Исчерченность
Характерная форма и размеры политенных хромосом достигаются вследствие их максимальной деспирализации и многократного воспроизведения хромосом без их последующего расхождения, то есть они образуются как результат эндомитоза .
Политенные хромосомы имеют характерную поперечную исчерченность, обусловленную наличием участков более плотной спирализации хромонем - хромомер . В тёмных участках (то есть хромомерах) располагается спирализованный неактивный хроматин , в то время как светлые полосы указывают на участок с повышенной транскрипционной активностью. Чёткое различение тёмных дисков и светлых междисковых участков объясняется нерасхождением дочерних хромонем. По этой причине все особенности отдельной хромонемы, в том числе хромомерный рисунок, становятся выраженными более контрастно .
По сути, политенные хромосомы представляют собой пару гигантских гомологичных хромосом , находящихся в состоянии идеально точной соматической конъюгации . При этом диски и междисковые участки гомологов расположены строго параллельно и тесно сближены. Такая конъюгация не характерна для подавляющего большинства соматических клеток .
Впервые карта политенных хромосом была составлена в 1935 году Кэлвином Бриджесом , и она широко используется и по сей день.
Уникальность строения политенных хромосом, а именно возможность чётко различать детали их строения, была использована Т. Пайтнером для изучения их перестроек и характера конъюгации . Вообще, исчерченность политенных хромосом исключительно полезна для исследований, в частности, на примере политенных хромосом была получена визуализация участков активного и неактивного хроматина. На них также можно изучать общую структуру хроматина.
Кроме того, политенные хромосомы помогают идентифицировать личинок комаров-звонцов (Chironomid ), которых другим способом отличить сложно .
Пуфы
В политенных хромосомах процесс транскрипции сопровождается формированием т. н. пуфов - характерных вздутий определённых дисков, образующихся в результате локальной декомпактизации в них ДНК . На активную транскрипцию в этих регионах указывает активное включение 3 H-уридина в районе пуфов. Крупные пуфы называются кольцами Бальбиани (в некоторых источниках термины «пуф» и «кольца Бальбиани» употребляют как синонимичные) .
Таким образом, образование пуфов является ярким примером дифференциальной транскрипции . Другим известным примером этого процесса является формирование хромосом типа ламповых щёток .
Функции
Политенные хромосомы содержат большое число копий генов, что многократно усиливает генную экспрессию . Это, в свою очередь, увеличивает производство необходимых белков. Например, в клетках слюнных желёз личинок D. melanogaster политенизация хромосом необходима для образования большого количества клейкого вещества до окукливания .
Примечания
- , с. 66-70.
- , с. 69.
- Balbiani E. G. Sur la structure du noyau des cellules salivaires chez les larves de Chironomus (фр.) // Zoologischer Anzeiger (англ.) русск. : magazine. - 1881. - Vol. 4 . - P. 637-641 .
КОНЪЮГАЦИЯ ХРОМОСОМ (лат. conjugatio соединение; хромосомы; син.: синапсис хромосом, спаривание хромосом ) - тесное соединение хромосом друг с другом у всех организмов, включая человека, обладающих сформированным клеточным ядром.
Различают конъюгацию гомологичных и негомологичных хромосом. Конъюгация гомологичных хромосом является обязательным этапом мейоза (см.), а также происходит в некоторых соматических клетках, напр, при формировании политенных (гигантских) хромосом (см.) в клетках слюнных желез личинок мух, комаров и других двукрылых насекомых. Этот тип К. х. отличается тем, что гомологичные хромосомы за счет специфического взаимного «узнавания» и притяжения гомологичных генов плотно прилегают одна к другой по всей длине так, что хромомеры, содержащие гомологичные (аллельные) гены, находятся точно друг против друга. Если в одной из конъюгирующих хромосом произошла транслокация (см.) или инверсия (см.), то участок хромосомы с измененной последовательностью расположения генов не способен конъюгировать с противолежащим участком второй, гомологичной хромосомы (рис. 1). Однако если перестройка произошла на достаточно длинном отрезке, то хромосомы-партнеры, образуя петлю или крест, способны обеспечить сопоставление гомологичных локусов и тем самым осуществить конъюгацию.
В политенных хромосомах конъюгация гомологов носит необратимый характер, гомологичные хромосомы остаются соединенными до конца существования слюнных желез. Биол. значение такой К. х. остается неясным. Конъюгация гомологичных хромосом в мейозе носит обратимый характер и лежит в основе точного разделения диплоидного набора хромосом на два гаплоидных набора, которые расходятся в разные клетки (редукция числа хромосом). Это явление создает условия для полового процесса и генетической рекомбинации у диплоидных организмов. Процесс конъюгации гомологичных хромосом происходит В профазе I мейотического деления и начинается на стадии зиготены. При этом сначала происходит сближение гомологичных хромосом с расстояния нескольких микрометров до расстояния примерно 0,2 мкм, обеспечивающего контакт хромосом. Затем происходит собственно «узнавание» и специфическое притяжение гомологичных хромомеров (локусов генов). В результате две конъюгирующие гомологичные хромосомы ложатся параллельно одна другой так, что гомологичные хромомеры образуют пары. Две соединенные гомологичные хромосомы носят название бивалента, а стадия мейоза, на к-рой соединение (конъюгация) завершилось по всей длине хромосом, называется пахитеной. Во время пахитены происходит кроссинговер (перекрест) - обмен участками гомологичных хромосом (см. Рекомбинация). На стадии пахитены каждая хромосома состоит из двух продольных половин - хроматид (4 хроматиды в биваленте). В данном месте бивалента кроссинговер происходит только между двумя несестринскими хроматидами из четырех. На следующей стадии - в диплотене - гомологичные хромосомы отталкиваются одна от другой во всех точках бивалента, кроме тех, где произошли перекресты. В результате места перекрестов (хиазмы) становятся видимыми в микроскоп. На следующих стадиях мейоза - в диплотене, диакинезе и метафазе I деления под влиянием конденсации и укорочения хромосом хиазмы перемещаются к концам бивалентов. Происходит так наз. терминализация хиазм. При этом хиазмы продолжают удерживать пары бывших партнеров по конъюгации. В метафазе I каждая хромосома в биваленте соединяется нитью веретена только с одним полюсом клеточного деления. Благодаря этому в анафазе I мейотического деления гомологичные хромосомы расходятся к противоположным полюсам и к каждому полюсу попадает по одной хромосоме из каждого бивалента. Таким образом, К. х. и хиазмы обеспечивают правильную редукцию числа хромосом. Если в кариотипе (см.) есть непарные хромосомы, напр, единичная половая хромосома в норме у самцов некоторых видов насекомых или одиночная половая хромосома у человека при синдроме Тернера, то такие хромосомы не вступают в конъюгацию из-за отсутствия партнера, остаются унивалентными в профазе I и случайным образом направляются к тому или иному полюсу в анафазе I. У противоположного полюса оказывается набор, лишенный одной хромосомы. У межвидовых гибридов, напр, у мула, нет ни одной парной хромосомы, т. к. половина хромосом получена от хромосомного набора лошади, а другая - осла. В результате в профазе I К. х. вообще отсутствует, и все хромосомы остаются унивалентными. В анафазе I деления они хаотически расходятся к полюсам, и дочерние клетки, а также формирующиеся из них гаметы получают несбалансированные как по видовому составу, так и по числу наборы хромосом. Это приводит к нежизнеспособности гамет или зигот. Бесплодие гибридов типа мула обусловлено отсутствием у них К. х. в мейозе.
Появление в диплоидном хромосомном наборе (см.) лишней хромосомы (трисомия по данной хромосоме) приводит к нарушению К. х. из-за конкуренции между тремя партнерами по конъюгации. Исследование таких случаев привело к открытию правила, согласно к-рому в каждой точке (локусе) хромосомы возможна конъюгация только двух партнеров. Однако в другом локусе может произойти смена партнера, и в результате появляются триваленты. Это также нарушает расхождение хромосом в анафазе I. Различные случаи нарушения К. х. приводят к появлению гипогаплоидных или гипергаплоидных гамет (недостаток хромосом или их избыток). Если такие гаметы выживают и формируют зиготу, то возникают анэуплоидные зародыши (нарушение строгой диплоидности), при этом возможны случаи моносомии (отсутствие одной из хромосом), трисомии (появление лишней хромосомы) и т. п. У человека моно- и трисомия по половым хромосомам и нек-рым аутосомам приводит к хромосомным болезням (см.), а анэуплоидия по крупным аутосомам - к гибели плода во внутриутробном периоде.
Иногда у гибридов близких видов, в частности у растительных гибридов, или у организмов, несущих крупные перестройки хромосом, наблюдается частичная гомология хромосом. Тогда о способности хромосом к конъюгации судят по числу бивалентов в клетке на стадиях диплотены - метафазы I и о «силе» этой конъюгации - по числу хиазм на бивалент. Однако известны случаи ахиазматии (полного отсутствия хиазм) при нормально протекающей конъюгации хромосом в зиготене и пахитене. Преждевременное разъединение гомологов (распад бивалентов) из-за отсутствия хиазм называют десинапсисом. Десинапсис приводит к такому же нарушению расхождения хромосом в анафазе I, какое наблюдается у мула.

Установлено, что процесс конъюгации гомологичных хромосом в мейозе находится под контролем многих генов, действующих только во время мейоза. Такие гены обнаружены у дрозофилы, грибов и ряда высших растений, но их действие оказывается одинаковым у самых разных организмов. Генетический контроль К. х. свидетельствует о том, что К. х. обеспечивается синтезируемыми во время мейоза специфическими белками. Сближение хромосом с дальних расстояний, вероятно, осуществляется за счет каких-то факторов в ядерной мембране: концы гомологичных хромосом, прикрепленные к ядерной мембране, «скользят» по ней навстречу друг другу, обеспечивая тем самым сближение хромосом. Возможно, что сближение хромосом с дальних расстояний происходит за счет неспецифического взаимодействия ДНК, локализованной в прицентромерных участках хромосом, за счет интеркалярного гетерохроматина (т. е. гетерохроматина, находящегося между двумя дисками эухроматина), способного к так наз. эктопическому спариванию - временной конъюгации негомологичных участков хромосом. Более известны молекулярные механизмы взаимодействия хромосом на близких расстояниях. Доказано участие в этом процессе уникальных последовательностей нуклеотидов ДНК, расположенных по ли локально по длине всех хромосом и реплицирующихся накануне или во время К. х. на стадии зиготены (зетДНК). Установлено, что К. х. на стадии зиготены сопровождается формированием так наз. синаптонемального комплекса (СК). Он формируется в процессе К. х. в мейозе у всех эукариотических организмов (от инфузорий и дрожжей до человека) и представляет собой субмикроскопическую структуру, к-рая состоит из трех рибонуклеопротеидных тяжей, идущих вдоль каждой пары гомологичных хромосом внутри бивалента (рис. 2). Два наружных тяжа называются боковыми элементами СК, внутренний - центральным. Боковые элементы возникают в каждой хромосоме до их конъюгации и сближаются в момент конъюгации до расстояния 150-200 нм. В это время между ними формируется центральный элемент. Предполагается, что центральный элемент служит местом формирования гетеро-дуплексов зетДНК (гибридных молекул ДНК), в к-рой полинуклеотидные нити двойной спирали принадлежат разным хромосомам - партнерам в биваленте. Предполагается, что СК препятствует необратимому соединению гомологичных хромосом, удерживая их на строго определенном расстоянии, ибо он отторгается от хромосом после прекращения их конъюгации на стадии диплотены, разрушается и выводится из ядра нередко в виде поликомплексов, упакованных в трехмерный штабель отрезков боковых и центральных элементов. СК отсутствует при необратимой К. х. в политенных хромосомах. Доказано, что формирование СК является результатом активации в мейозе специфических генов, в частности нормального аллеля гена с (3) G у дрозофилы. Формирование СК обеспечивает высокую частоту кроссинговера, но не является непременным условием для его осуществления; в отсутствие СК кроссинговер может идти, но с пониженной частотой.
Конъюгация негомологичных хромосом (конъюгация гомологичных участков в негомологичных хромосомах) наблюдается в мейозе у гаплоидных растений, во время митоза (см.) в соматических клетках многих растений и животных (эктопическое спаривание). На основе неслучайного расхождения негомологичных хромосом в мейозе у дрозофил с перестроенным хромосомным набором можно сделать вывод, что в профазе I мейоза происходила конъюгация негомологичных хромосом. Экспериментально подтверждается гипотеза о полилокальном расположении в хромосомах дрозофилы определенной фракции ДНК, состоящей из так наз. умеренных повторов нуклеотидов. Они способны обеспечить взаимное «узнавание» идентичных участков в той же самой и других хромосомах клетки и обусловить, т. о., конъюгацию негомологичных хромосом.
Библиография: Дубинин Н. П. Общая генетика, М., 1976; ДыбанА. П. иБара-н о в В. С. Методы исследования хромосом в гаметогенезе и эмбриогенезе млекопитающих, Арх. анат., гистол, и эмбриол., т. 66, № 1, с. 79, 1974, библиогр.; Прокофьева - Бельговская А. А. и д р. Цитогенетика человека, М., 1969; Цитология и генетика мейоза, под ред. В. В. Хвостовой и Ю. Ф. Богданова, М., 1975, библиогр.; Bordjadze V. К. a. Prokofieva-Belgov-skaya A. A. Pachytene analysis of human acrocentric chromosomes, Cytogenetics, v. 10, p. 38, 1971; J o h n B. a. L e w i s K. K. The meiotic system, Wien-N. Y., 1965, bibliogr.; Methods in human cytogenetics, ed. by H. G. Schwarzacher a. U. Wolf, N. Y., 1974.
Ю. Ф. Богданов.
В два раза. Происходит в два этапа (редукционный и эквационный этапы мейоза). Мейоз не следует смешивать с гаметогенезом - образованием специализированных половых клеток , или гамет , из недифференцированных стволовых .
С уменьшением числа хромосом в результате мейоза в жизненном цикле происходит переход от диплоидной фазы к гаплоидной. Восстановление плоидности (переход от гаплоидной фазы к диплоидной) происходит в результате полового процесса .
В связи с тем, что в профазе первого, редукционного, этапа происходит попарное слияние (конъюгация) гомологичных хромосом, правильное протекание мейоза возможно только в диплоидных клетках или в чётных полиплоидах (тетра-, гексаплоидных и т. п. клетках). Мейоз может происходить и в нечётных полиплоидах (три-, пентаплоидных и т. п. клетках), но в них, из-за невозможности обеспечить попарное слияние хромосом в профазе I, расхождение хромосом происходит с нарушениями, которые ставят под угрозу жизнеспособность клетки или развивающегося из неё многоклеточного гаплоидного организма.
Этот же механизм лежит в основе стерильности межвидовых гибридов . Поскольку у межвидовых гибридов в ядре клеток сочетаются хромосомы родителей, относящихся к различным видам, хромосомы обычно не могут вступить в конъюгацию. Это приводит к нарушениям в расхождении хромосом при мейозе и, в конечном счете, к нежизнеспособности половых клеток, или гамет . Определенные ограничения на конъюгацию хромосом накладывают и хромосомные мутации (масштабные делеции, дупликации, инверсии или транслокации).
Фазы мейоза
Мейоз состоит из двух последовательных делений с короткой интерфазой между ними.
- Профаза I - профаза первого деления очень сложная и состоит из 5 стадий:
- Фаза лептотены или лептонемы - конденсация ДНК с образованием хромосом в виде тонких нитей.
- Зиготена или зигонема - коньюгация (соединение) гомологичных хромосом с образованием структур, состоящих из двух соединённых хромосом, называемых тетрадами или бивалентами.
- Пахитена или пахинема - кроссинговер (перекрест) обмен участками между гомологичными хромосомами; гомологичные хромосомы остаются соединенными между собой.
- Диплотена или диплонема - происходит частичная деконденсация хромосом, при этом часть генома может работать, происходят процессы транскрипции (образование РНК), трансляции (синтез белка); гомологичные хромосомы остаются соединёнными между собой.
- Диакинез - ДНК снова максимально конденсируется, синтетические процессы прекращаются, растворяется ядерная оболочка; гомологичные хромосомы остаются соединёнными между собой.
- Метафаза I - бивалентные хромосомы выстраиваются вдоль экватора клетки.
- Анафаза I - микротрубочки сокращаются, биваленты делятся и хромосомы расходятся к полюсам. Важно отметить, что, из-за конъюгации хромосом в зиготене, к полюсам расходятся целые хромосомы, состоящие из двух хроматид каждая, а не отдельные хроматиды, как в митозе .
- Телофаза I
Второе деление мейоза следует непосредственно за первым, без выраженной интерфазы: S-период отсутствует, поскольку перед вторым делением не происходит репликации ДНК.
- Профаза II - происходит конденсация хромосом, клеточный центр делится и продукты его деления расходятся к полюсам ядра, разрушается ядерная оболочка, образуется веретено деления.
- Метафаза II - унивалентные хромосомы (состоящие из двух хроматид каждая) располагаются на «экваторе» (на равном расстоянии от «полюсов» ядра) в одной плоскости, образуя так называемую метафазную пластинку.
- Анафаза II - униваленты делятся и хроматиды расходятся к полюсам.
- Телофаза II - хромосомы деспирализуются и появляется ядерная оболочка.