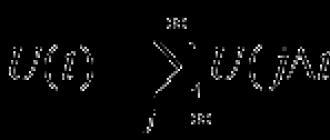Биологическое окисление: суть процесса и его виды. Биологическое окисление Где происходит процесс биологического окисления
Осуществление биосинтеза биоло-гических молекул требует постоянного притока энергии. В автотрофных кле-точных биосистемах такой энергией служит лучистая энергия Солнца. В гетеротрофных клеточных биосистемах процесс биосинтеза идёт с помо-щью энергии, получаемой от расщепления молекул органических веществ — уг-леводов , белков и жиров . Регулярное поступление энергии является необходи-мым условием биосинтеза. Поэтому синтез органических веществ сопровож-дается непрерывной доставкой требуемой энергии, освобождающейся при расщеплении молекул других органических соединений, имеющихся в клетке .
Процесс высвобождения необходимой энергии из органических ве-ществ путём их расщепления называют биологическим окислением или кле-точным дыханием .
При этом идёт накопление энергии в виде молекул АТФ и других макроэнергетических соединений.
АТФ , или аденозинтрифосфорная кислота , — это нуклеотид, состоя-щий из аденина, рибозы и трёх остатков фосфорной кислоты — трифосфата (рис. 66).
Окисление осуществляется путём отрыва от окисляемой молекулы электронов или атома водорода. Такая потеря всегда сопровождается выделени-ем значительного количества энергии. Это объясняется тем, что электроны в соста-ве молекул органических соединений находятся на очень высоких энергетических уровнях этих молекул. Перемещаясь с высоких на более низкие уровни своей или другой молекулы или атома, электроны высвобождают энергию. Молекулы, те-ряющие электроны, называются донорами, а принимающие их — акцепторами. Конечным акцептором в окислительных процессах клетки часто выступает кисло-род. Именно поэтому кислород так важен для дыхания многим организмам . Окан-чивается биологическое окисление (клеточ-ное дыхание) органических соединений образованием воды и углекислого газа. На примере окисления глюкозы этот процесс клеточного дыхания выражается обобщён-ным уравнением:
C 6 H 12 O 6 + 6O 2 → 6CO 2 + 6H 2 O + Энергия;
Для осуществления клеточного дыхания многим организмам нужен кислород, в этом случае говорят об аэробном (кислородном) дыхании или аэробном способе высвобожде-ния энергии. Однако в природе существует множество различных групп организмов, которые могут получать энергию для своей жизнедеятельности без использования свободного атмо-сферного кислорода, т. е. путём бескислородного или анаэробного клеточно-го дыхания (анаэробное высвобождение энергии).
Основным веществом, используемым для получения энергии, в клетке обычно служат жиры и глюкоза. Процессы окисления глюкозы протекают в не-сколько этапов и сопровождаются ступенчатым выделением энергии, что обеспечивает возможность её запасания и дальнейшего перехода в макроэнергетическую связь в виде молекул аденозинтрифосфорной кислоты — АТФ.
Молекула АТФ необычайно энергоемка. Это связано с тем, что её трифосфатный компонент содержит две фосфорангидридные связи. Разрыв од-ной из них, т. е. отделение от АТФ одного концевого фосфата (Ф Н), сопро-вождается выделением 40 кДж на 1 моль вместо 12 кДж, выделяемых при разрыве обычных химических связей других соединений. Образовавшаяся при этом молекула аденозиндифосфата (АДФ) с двумя фосфатными остатками может быстро восстановиться до АТФ или, при необходимости, отдать ещё один концевой фосфат и превратиться в адепозинмонофосфат (АМФ). Упро-щенно процессы можно представить на схемах освобождения энергии при отделении концевых фосфатов у АТФ и АДФ (1) до АМФ, а также восстановле-ния АТФ из АДФ (2):
 |
Высвобождение энергии из химических связей органических веществ в общих чертах напоминает обратную последовательность процессов её свя-зывания. Поэтому можно сказать, что клеточное дыхание противоположно фотосинтезу и в схематическом виде оно распадается на два этапа: бескисло-родный этап (гликолиз) и кислородный этап. Материал с сайта
В отличие от гликолиза, кисло-родный этап клеточного дыхания является мембранозависимым. Он осу-ществляется в матриксе митохондрий и на мембранах их крист. Здесь происходит полное окисление пирувата до конечных продуктов — CO 2 и H 2 O. В связи с этим различают две стадии, связанные с окислительным циклом трикарбоновых кислот (цикл Кребса, или лимонный цикл) и с дыхательной цепью переноса электронов , где синтезируется АТФ.
Наличие в живой материи разных высокоупорядоченных биохимиче-ских процессов гликолиза (в цитоплазме), кислородного дыхания (в мито-хондриях), как и фотосинтеза (в хлоропластах), позволяет судить о наличии в живой клетке целостных молекулярных биологических систем, а также о молекулярном структурном уровне как исключительно важном, основопола-гающем и обеспечивающем все процессы жизни на Земле.
На этой странице материал по темам:
Биологическое окисление клеточное дыхание
Кислородное окисление или дыхание
Биологическое окисление биохимия кратко
В чем биологическая роль окисления
Доклад по клеточному дыханию
Вопросы по этому материалу:
Без энергии невозможно существование ни одного живого существа. Ведь каждая химическая реакция, любой процесс требуют ее присутствия. Любому человеку легко понять это и почувствовать. Если весь день не употреблять пищу, то уже к вечеру, а возможно, и раньше, начнутся симптомы повышенной усталости, вялости, сила значительно уменьшится.
Каким же способом разные организмы приспособились к получению энергии? Откуда она берется и какие процессы при этом происходят внутри клетки? Попробуем разобраться в данной статье.
Получение энергии организмами
Каким бы способом ни потребляли существа энергию, в основе всегда лежат Примеры можно привести разные. Уравнение фотосинтеза, который осуществляют зеленые растения и некоторые бактерии − это тоже ОВР. Естественно, что процессы будут отличаться в зависимости от того, какое живое существо имеется в виду.
Так, все животные − это гетеротрофы. То есть такие организмы, которые не способны самостоятельно формировать внутри себя готовые органические соединения для дальнейшего их расщепления и высвобождения энергии химических связей.
Растения, напротив, являются самым мощным продуцентом органики на нашей планете. Именно они осуществляют сложный и важный процесс под названием фотосинтез, который заключается в формировании глюкозы из воды, углекислого газа под действием специального вещества − хлорофилла. Побочным продуктом является кислород, который является источником жизни для всех аэробных живых существ.
Окислительно-восстановительные реакции, примеры которых иллюстрируют данный процесс:
- 6CO 2 + 6H 2 O = хлорофилл = C 6 H 10 O 6 + 6O 2 ;
- диоксид углерода + под воздействием пигмента хлорофилла (фермент реакции) = моносахарид + свободный молекулярный кислород.
Также существуют и такие представители биомассы планеты, которые способны использовать энергию химических связей неорганических соединений. Их называют хемотрофы. К ним относят многие виды бактерий. Например, водородные микроорганизмы, окисляющие молекулы субстрата в почве. Процесс происходит по формуле: 2Н 2 +0 2 =2Н 2 0.

История развития знаний о биологическом окислении
Процесс, который лежит в основе получения энергии, сегодня вполне известен. окисление. Биохимия настолько подробно изучила тонкости и механизмы всех стадий действия, что загадок почти не осталось. Однако так было не всегда.
Первые упоминания о том, что внутри живых существ происходят сложнейшие преобразования, которые являются по природе химическими реакциями, появились примерно в XVIII веке. Именно в это время Антуан Лавуазье, знаменитый французский химик, обратил свое внимание на то, как схожи биологическое окисление и горение. Он проследил примерный путь поглощаемого при дыхании кислорода и пришел к выводу, что внутри организма происходят процессы окисления, только более медленные, чем снаружи при горении различных веществ. То есть окислитель − молекулы кислорода − вступают в реакцию с органическими соединениями, а конкретно, с водородом и углеродом из них, и происходит полное превращение, сопровождающееся разложением соединений.
Однако, хоть данное предположение по сути своей вполне реально, непонятными оставались многие вещи. Например:
- раз процессы схожи, то и условия их протекания должны быть идентичными, но окисление происходит при низкой температуре тела;
- действие не сопровождается выбросом колоссального количества тепловой энергии и не происходит образования пламени;
- в живых существах не менее 75-80% воды, но это не мешает «горению» питательных веществ в них.
Чтобы ответить на все эти вопросы и понять, что на самом деле представляет собой биологическое окисление, понадобился не один год.
Существовали разные теории, которые подразумевали важность наличия в процессе кислорода и водорода. Самые распространенные и наиболее успешные были:
- теория Баха, именуемая перекисной;
- теория Палладина, основывающаяся на таком понятии, как «хромогены».
В дальнейшем было еще много ученых, как в России, так и других странах мира, которые постепенно вносили дополнения и изменения в вопрос о том, что же такое биологическое окисление. Биохимия современности, благодаря их трудам, может рассказать о каждой реакции этого процесса. Среди самых известных имен в этой области можно назвать следующие:
- Митчелл;
- С. В. Северин;
- Варбург;
- В. А. Белицер;
- Ленинджер;
- В. П. Скулачев;
- Кребс;
- Грин;
- В. А. Энгельгардт;
- Кейлин и другие.

Виды биологического окисления
Можно выделить два основных типа рассматриваемого процесса, которые протекают при разных условиях. Так, самый распространенный у многих видов микроорганизмов и грибков способ преобразования получаемой пищи − анаэробный. Это биологическое окисление, которое осуществляется без доступа кислорода и без его участия в какой-либо форме. Подобные условия создаются там, куда нет доступа воздуху: под землей, в гниющих субстратах, илах, глинах, болотах и даже в космосе.
Этот вид окисления имеет и другое название − гликолиз. Он же является одной из стадий более сложного и трудоемкого, но энергетически богатого процесса − аэробного преобразования или тканевого дыхания. Это уже второй тип рассматриваемого процесса. Он происходит во всех аэробных живых существах-гетеротрофах, которые для дыхания используют кислород.
Таким образом, виды биологического окисления следующие.
- Гликолиз, анаэробный путь. Не требует присутствия кислорода и заканчивается разными формами брожения.
- Тканевое дыхание (окислительное фосфорилирование), или аэробный вид. Требует обязательного наличия молекулярного кислорода.

Участники процесса
Перейдем к рассмотрению непосредственно самих особенностей, которые заключает в себе биологическое окисление. Определим основные соединения и их аббревиатуры, которые в дальнейшем будем использовать.
- Ацетилкоэнзим-А (ацетил-КоА) − конденсат щавелевой и уксусной кислоты с коферментом, формирующийся на первой стадии цикла трикарбоновых кислот.
- Цикл Кребса (цикл лимонной кислоты, трикарбоновых кислот) − ряд сложных последовательных окислительно-восстановительных преобразований, сопровождающихся высвобождением энергии, восстановлением водорода, образованием важных низкомолекулярных продуктов. Является главным звеном ката- и анаболизма.
- НАД и НАД*Н − фермент-дегидрогеназа, расшифровывающийся как никотинамидадениндинуклеотид. Вторая формула − это молекула с присоединенным водородом. НАДФ - никотинамидадениндинуклетид-фосфат.
- ФАД и ФАД*Н − флавинадениндинуклеотид - кофермент дегидрогеназ.
- АТФ − аденозинтрифосфорная кислота.
- ПВК − пировиноградная кислота или пируват.
- Сукцинат или янтарная кислота, Н 3 РО 4 − фосфорная кислота.
- ГТФ − гуанозинтрифосфат, класс пуриновых нуклеотидов.
- ЭТЦ − электроно-транспортная цепь.
- Ферменты процесса: пероксидазы, оксигеназы, цитохромоксидазы, флавиновые дегидрогеназы, различные коферменты и прочие соединения.
Все эти соединения являются непосредственными участниками процесса окисления, которое происходит в тканях (клетках) живых организмов.
Стадии биологического окисления: таблица
| Стадия | Процессы и значение |
| Гликолиз | Суть процесса заключается в бескислородном расщеплении моносахаридов, которое предшествует процессу клеточного дыхания и сопровождается выходом энергии, равным двум молекулам АТФ. Также образуется пируват. Это начальная стадия для любого живого организма гетеротрофа. Значение в образовании ПВК, который поступает на кристы митохондрий и является субстратом для тканевого окисления кислородным путем. У анаэробов после гликолиза наступают процессы брожения разного типа. |
| Окисление пирувата | Этот процесс заключается в преобразовании ПВК, образовавшейся в ходе гликолиза, в ацетил-КоА. Он осуществляется при помощи специализированного ферментного комплекса пируватдегидрогеназы. Результат − молекулы цетил-КоА, которые вступают в В этом же процессе осуществляется восстановление НАД до НАДН. Место локализации − кристы митохондрий. |
| Распад бета-жирных кислот | Этот процесс осуществляется параллельно с предыдущим на кристах митохондрий. Суть его в том, чтобы переработать все жирные кислоты в ацетил-КоА и поставить его в цикл трикарбоновых кислот. При этом также восстанавливается НАДН. |
| Цикл Кребса | Начинается с превращения ацетил-КоА в лимонную кислоту, которая и подвергается дальнейшим преобразованиям. Одна из важнейших стадий, которые включает в себя биологическое окисление. Данная кислота подвергается:
Каждый процесс совершается несколько раз. Результат: ГТФ, диоксид углерода, восстановленная форма НАДН и ФАДН 2 . При этом ферменты биологического окисления свободно располагаются в матриксе митохондриальных частиц. |
| Окислительное фосфорилирование | Это последняя стадия преобразования соединений в организмах эукариот. При этом происходит преобразование аденозиндифосфата в АТФ. Энергия, необходимая для этого, берется при окислении тех молекул НАДН и ФАДН 2 , которые сформировались на предыдущих стадиях. Путем последовательных переходов по ЭТЦ и понижением потенциалов происходит заключение энергии в макроэргические связи АТФ. |
Это все процессы, которые сопровождают биологическое окисление при участии кислорода. Естественно, что описаны они не полностью, а лишь по сущности, так как для подробного описания нужна целая глава книги. Все биохимические процессы живых организмов чрезвычайно многогранны и сложны.

Окислительно-восстановительные реакции процесса
Окислительно-восстановительные реакции, примеры которых могут проиллюстрировать описанные выше процессы окисления субстрата, следующие.
- Гликолиз: моносахарид (глюкоза) + 2НАД + + 2АДФ = 2ПВК + 2АТФ + 4Н + + 2Н 2 О + НАДН.
- Окисление пирувата: ПВК + фермент = диоксид углерода + ацетальдегид. Затем следующий этап: ацетальдегид + Кофермент А = ацетил-КоА.
- Множество последовательных преобразований лимонной кислоты в цикле Кребса.
Данные окислительно-восстановительные реакции, примеры которых приведены выше, отражают суть происходящих процессов лишь в общем виде. Известно, что соединения, о которых идет речь, относятся к высокомолекулярным, либо имеющим большой углеродный скелет, поэтому изобразить все полными формулами просто не представляется возможным.
Энергетический выход тканевого дыхания
По приведенным выше описаниям очевидно, что подсчитать суммарный выход всего окисления по энергии несложно.
- Две молекулы АТФ дает гликолиз.
- Окисление пирувата 12 молекул АТФ.
- 22 молекулы приходится на цикл трикарбоновых кислот.
Итог: полное биологическое окисление по аэробному пути дает выход энергии, равный 36 молекулам АТФ. Значение биологического окисления очевидно. Именно эта энергия используется живыми организмами для жизни и функционирования, а также для согревания своего тела, движения и прочих необходимых вещей.

Анаэробное окисление субстрата
Второй вид биологического окисления − анаэробный. То есть тот, что осуществляется у всех, но на котором останавливаются микроорганизмы определенных видов. и именно с него четко прослеживаются различия в дальнейшем преобразовании веществ между аэробами и анаэробами.
Стадии биологического окисления по данному пути немногочисленны.
- Гликолиз, то есть окисление молекулы глюкозы до пирувата.
- Брожение, приводящее к регенерации АТФ.
Брожение может быть разных типов, в зависимости от организмов, его осуществляющих.
Молочнокислое брожение
Осуществляется молочнокислыми бактериями, а также некоторыми грибками. Суть состоит в восстановлении ПВК до молочной кислоты. Этот процесс используют в промышленности для получения:
- кисломолочных продуктов;
- квашеных овощей и фруктов;
- силоса для животных.
Этот вид брожения является одним из самых применяемых в нуждах человека.
Спиртовое брожение
Известно людям с самой древности. Суть процесса заключается в превращении ПВК в две молекулы этанола и две диоксида углерода. Благодаря такому выходу продукта, данный вид брожения используют для получения:
- хлеба;
- вина;
- пива;
- кондитерских изделий и прочего.
Осуществляют его грибы дрожжи и микроорганизмы бактериальной природы.

Маслянокислое брожение
Достаточно узкоспецифичный вид брожения. Осуществляется бактериями рода Клостридиум. Суть состоит в превращении пирувата в масляную кислоту, придающую продуктам питания неприятный запах и прогорклый вкус.
Поэтому реакции биологического окисления, идущие по такому пути, практически не используют в промышленности. Однако эти бактерии самостоятельно засевают продукты питания и наносят вред, понижая их качество.
Биоэнергетические процессы, приводящие к синтезу АТФ, к зарядке «биологических аккумуляторов», протекают в специализированных мембранах митохондрий. Именно здесь локализованы и пространственно организованы молекулярные системы, ответственные за энергетику живых организмов. Синтез АТФ в митохондриях сопряжен с электронным и ионным транспортом и с механохимическими явлениями. Функции митохондриальных мембран весьма сложны и многообразны. Другой тип биоэнергетических сопрягающих мембран - мембраны хлоропластов растений, ответственные за фотосинтез, - рассматривается в гл.7.
Источником энергии, расходуемой клеткой на биосинтез, активный транспорт, механическую и электрическую работу, является дыхание, т. е. окисление органических соединений кислородом воздуха. В 1780 г. Лавуазье показал, что дыхание и горение имеют единую природу. За последующие почти два столетия исследования химиков, биологов и физиков привели к раскрытию основных особенностей биологического окисления - одного из важнейших процессов (или, скорее, системы процессов), происходящих в живой природе.
Топливо, т. е. окисляемые вещества, поступает в организм животного с пищей в виде жиров, углеводов и белков. Жиры представляют собой триглицериды жирных кислот, главным образом многоатомных. Они расщепляются, т. е. гидролизуются в реакциях, катализируемых специальными ферментами. Жирные кислоты активируются с участием специфических ферментов и АТФ, превращаясь в ацилпроизводные так называемого кофер - мента А, Ко А - SH, структура которого показана на рис. 6.1. Окисление ацилпроизводного Ко А - SH происходит в ряде стадий, на каждой из которых образуется остаток жирной кислоты, содержащий на два атома углерода меньше, чем предыдущий . Полное уравнение реакции окисления жирной кислоты с четным числом атомов углерода до ацетил-S - Ко А имеет вид
Н3С(СН2СН2)„С02Н + АТФ + (п + 1) КоА - SH + лНАД+ +
ПЕ ■ ФАД + лН20 ->- (п + 1)CH3COS - КоА +
+ (£5ф + £фч) + "НАД-Н + пЕ - ФАД-Н +
Здесь НАД - кофермент никотинамидадениндинуклеотид (рис.
6.2), Е - фермент, ФАД- кофермент флавинадениндинуклеотид
Рис. 6.1. Кофермент А.
TOC \o "1-3" \h \z (рис. 6.3), АДФ и АМФ - аденозиндифосфат и аденозинмоно - фосфат, Фн, ФФН - неорганические моно - и дифосфаты. Обратим внимание попутно на общность и
Строения АТФ, КоА-SH, НАД и Н В
ФАД (см. , гл. 2). При разруше - /\ / \
Нии жиров в конечном счете обра - н| 9 NH2
Зуется ацетил-КоА, а также про - п ^
ПИОНИЛ-КО А И ГЛИЦерИН. - g_p_Q_QH
Расщепление и окисление угле - і | |
Водов (в частности, крахмала) при - \ С^н н^С водит к образованию триозофосфа - і |>с«исг I NH
Тов и пировиноградной кислоты I н L L н I 2
(пирувата). н°-рГ° 0Н 0Н „ /ч
При разрушении белков, наряду / С N
С индивидуальными аминокислота - \ НСч J I
Ми, используемыми в синтезе бел - СН2 0 \
Ков de novo, образуются ацетил - N
Ко А, оксалацетат, а-кетоглутарат, INc^c/I
Фумарат и сукцинат. Эти процессы н і і н
Метаболизма детально изучены в он ОН
Современной биохимии (см. ). Рис. в-2- Никотинамидаденин-
Основные продукты расщепле - динуклеотид (НАД),
Ния и окисления жиров, углеводов
И белков претерпевают дальнейшие превращения в циклической системе реакций, именуемой циклом лимонной кислоты или циклом Кребса. Эта система локализована в митохондриях. Цикл Кребса, «образно говоря, та главная ось, вокруг которой вертится метаболизм почти всех существующих клеток... Цикл
Кребса является «фокусом», в котором сходятся все метаболические пути» .
Цикл Кребса изображен на рис. 6.4. За один оборот цикла, состоящего из восьми реакций, отмеченных на рисунке цифрами в кружках, происходит деградация одной молекулы ацетил-КоА
Н2с-СН-СН-СН-СН,-О-Р-О-Р-О-СН, n-^Ч
A/\A/NH он он
Н, С V N С s Н II
Рис. 6.3. Флавииадениндинуклеотид (ФАД).
Или одной молекулы пиру вата до С02 и Н20, т. е. «сгорание» этих молекул. Соответствующие суммарные реакции имеют вид
CH, COS - КоА + З НАЦ+ + (ФАД) + ГДФ + Ф„ + 2 Н20 -
2 С02 + КоА - SH + З НАД-Н + (ФАД-Н) + ГТФ + ЗН+, Пируват" + КоА - SH + НАД+ -* CH3COS - КоА + НАД-Н+ + Н+ + С02.
(Скобки обозначают, что ФАД тесно связан с белком.)
От цикла Кребса идут пути многих биосиитетических реакций- пути синтеза углеводов, липидов, пуринов, пиримидииов и порфирннов. Синтез белков также связан с циклом, в котором создаются предшественники ряда аминокислот. Вместе с тем, как мы увидим, биологическое окисление является источником энергии, запасаемой в АТФ и необходимой для биосинтетических процессов.
В реакциях цикла возникают С02 и ионы Н+. Одновременно происходит восстановление коферментов НАД и ФАД. Для непрерывного и полного биологического окисления эти кофермеи - ты должны окисляться вновь. Окисление осуществляется совокупностью переносчиков электронов, образующих цепь переноса электронов (ЦПЭ), фиксированную в митохондриях. ЦПЭ обеспечивает следующие реакции:
З НАД-Н + 1,5 02 + ЗН+ з НАД+ + 3 Н20 - 3 52,4 ккал/моль,
(ФАД-Н) + 0,5 О, - (ФАД) + Н20 - 36,2 ккал/моль.
Реакция ацетил-КоА имеет вид
CH3COS - КоА + 2 02 -> 2 С02 + Н20 + КоА - SH - 215,2 ккал/моль.
ЦПЭ, иначе именуемая дыхательной цепью, представляет собой
Углеводы-*- CH-CDC07 п
ПируВат у Жирные
У v4*- "Ацетил-КоК С02 2Н ■
^ с=о (Т) г«
Фн Сущинил-Ш KoA_SH Рис. 6.4. Цикл Кребса.

Тре-"ЪА.-изоцитрац
А-кетоглутарапі
НО-СН С02 fiOj (2)
" СН2 Оксалацетат Цитрат
FQ) l-тлат \ Vz
Полиферментную систему, акцептирующую электроны из цикла Кребса и цикла окисления жирных кислот.
Электрон переносится по следующей цепи : Сукцинат - ФП3 1
Субстрат -*■ НАД -»- ФП0 ->■ Цитохром Ь ->- -*■ Цитохром С) -» Цитохром с -»- Цитохромы а + аз -*■ 02.
Флавопротеиды - специфические ферменты, содержащие флави- новые коферменты-ФАД (см. рис. 6.3) и флавинмононуклео - тид ФМН, рибофлавин-5"-фосфат. Цитохромы содержат группу гема, атом железа которого при работе цепи подвергается окислению и восстановлению:
Fe2+ Fe3+ - f e~.
Перенос электронов в приведенной цепи происходит слева направо, завершаясь восстановлением кислорода, который соединяется с водородом и образует воду. Освобождаемый при окислении электрон соединяется со следующим звеном цепи. Перенос электронов сопровождается изменением свободной энергии, так как электроны перемещаются по каскаду возрастающих окислительно-восстановительных потенциалов. Их значения приведены в табл. 6.1 .
Таблица 6.1
Окислительно-восстановительные потенциалы некоторых систем
Перенос электронов по дыхательной цепи сопряжен с запасанием энергии в макроэргических молекулах АТФ. Иными словами, освобождаемая свободная энергия конвертируется в химическую энергию АТФ. Происходит окислительное фосфорили - рование.
Это важнейшее явление было впервые открыто Энгельгард - том в 1930 г. . Белицер и Цыбакова подробно изучили стехиометрические соотношения между окислением и фосфори - лированием, провели первые определения коэффициента Фн О. т. е. отношения числа молекул этерифицированного неорганиче
ского фосфата к числу атомов поглощенного кислорода, и показали, что значение этого коэффициента не менее 2. В работе были даны термодинамические оценки, показавшие, что энергия переноса электронов на кислород достаточна для образования двух или большего числа молекул АТФ на один атом поглощенного кислорода. Калькар установил, что аэробное фосфо - рилирование сопряжено с дыханием и не зависит от гликолити - ческого фосфорилирования . Количественные соотношения были уточнены Очоа . Коэффициент Фн: О для окислительных реакций цикла Кребса и реакций с участием НАД равен 3. Ленинджер впервые установил, что процессы окислительного фосфорилирования локализованы именно в митохондриях .
Прямые стрелки показывают места вхождения электронов. ФП,. ФП, ...-флавопротеиды.
KoQ - кофермент Q.
10], и в работах его лаборатории были найдены узловые пункты дыхательной цепи, в которых происходит фосфорилирование. Указанное значение Фн: О следует из уравнения
НАД-Н + Н+ + З АДФ + 3 Ф„ + V2O2 - НАД+ + 4 Н20 + 3 АТФ. В этом уравнении суммируются экзергоническая реакция НАД-Н + Н+ + V2O2 -> НАД+ + Н20 + 52,7 ккал/моль
И эндергоническая реакция
3 АДФ + 3 Фн ->■ З АТФ + 3 Н20 - 21,9 ккал/моль.
Фосфорилирование АДФ -> АТФ происходит в трех узловых пунктах - в участке цепи НАД-Н - флавопротеид, в участке цитохром Ь цитохром сі и в участке цитохром с -* цитохромы а + а3.
Общая схема сопряжения окисления с фосфорилированием приведена на рис. 6.5.
Напишем вновь брутто-уравнение фосфорилирования

ПируВат^ Сущинагп
АДФ + H2P04" - f Н+ +=± АТФ + HgO - ДО,
Где AG - изменение свободной энергии. Имеем
TOC \o "1-3" \h \z [адф] Гн, ро:1 [н+] , >4
AG = AG" + RT ІП. (6.1)
AG0 - стандартное изменение свободной энергии в калориях, т. е. значение AG при рН 7,0, 25°С и концентрациях всех компонентов, равных 1,0 М. Как известно,
AG0 = - RT In К, (6.2)
где К - константа равновесия реакции. Для фосфорилирования AG0 = 7,3 ккал/моль. Наблюдаемое значение AG in vivo зависит от концентрации протонов по обе стороны мембраны и, следовательно, от мембранной разности потенциалов. AG зависит также от концентрации ионов Mg++. При изменении рН от 6,0 до 9,0 при = 10 мМ AG меняется от 6,17 до 9,29 ккал/моль (см. ).
Изменение свободной энергии при переносе двух электронных эквивалентов по ЦПЭ от НАД-Н к 02 определяется разностью окислительно-восстановительных потенциалов 0,82 - (-0,32) = = 1,14 В, т. е.
AG0 = z Д-ф = - 2 23,06 -1,14 ккал/моль = - 52,7 ккал/моль.
Это обеспечивает с избытком синтез 3 молей АТФ из АДФ и Фн. Эффективность процесса выражается величиной 21,9/52,7, т. е. примерно равна 40%.
Энергетический смысл дыхания состоит в синтезе АТФ. Энергия, аккумулированная в АТФ, используется клеткой для выполнения всех видов ее работы.
Раскрытие химизма биологического окисления - крупнейшее достижение биохимии. Здесь изложены лишь некоторые важнейшие сведения, подробное рассмотрение сложной биохимической системы окисления содержится в специальной литературе (см. ).
Особенность системы окислительного фосфорилирования, отличающая ее от ряда ферментативных реакций, протекающих в растворе, состоит в строгой пространственной локализации звеньев многоступенчатого процесса. Окислительное фосфорилиро - вание локализовано в митохондриях и непосредственно связано с транспортной и механохимической функциональностью их мембран. По-видимому, столь сложная система биохимических реакций принципиально требует пространственной неоднородности и не могла бы реализоваться в гомогенной среде.
Расшифровка кратко описанных здесь окислительно-восстановительных реакций была получена в результате применения тонких химических и физических методов. Здесь, в частности,
сыграли большую роль работы Чаиса, посвященные спектроскопии переносчиков электронов (НАД, ФП, цитохромов) в интакт - ных митохондриях. Эти переносчики обладают характерными полосами поглощения в видимой и близкой ультрафиолетовой областях спектра, и разностные спектры позволяют изучать кинетику их окисления и восстановления . Были применены различные способы удаления из митохондрий специфических ферментов н. тем самым, сохранения лишь определенных звеньев
Рис. 6.6. Распределение белков по комплексам ЦПЭ І, II, III, IV.
А, Ь, с. Сі, йі- >нтохромы, Си - медьсодержащие белкн, (Fe -be) -негемнновое железо, fs - сукцинатдегидрогеназа, iq-НАД Н-дегидрогеназа.
Процесса. Митохондрии подвергались расчленению, из субмито- хондриальных частиц выделялись комплексы дыхательных ферментов, свободные от структурных белков. Такие комплексы оказалось возможным очищать и детально исследовать. Были проведены успешные опыты по восстановлению ЦПЭ из выделенных препаратов и растворимых ферментов. Наконец, очень ценная информация была получена в опытах по ингибированию отдельных стадий процесса и по разобщению окислительного фосфорилирования и переноса электронов (см. § 6.5).
Можно считать установленным, что переносчики ЦПЭ сгруппированы в четыре комплекса, именуемые комплексами Грина (см. ). Соответствующая схема приведена на рис. 6.6. Молекулярный вес каждого комплекса равен примерно 3-Ю5 Он содержит около 64% белка и 36% липидов. Электрон-переносящий
комплекс определяется как минимальная единица ЦПЭ, сохраняющая способность переносить электрон со скоростью, сравнимой с этой скоростью в интактных митохондриях.
Изучение биологического окисления встречается с необходимостью решения ряда физических проблем, относящихся к различным уровням организации и функционирования системы.
Структура и функции молекулярных переносчиков электронов изучены еще недостаточно. В § 6.7 рассмотрены современные данные, относящиеся к строению и свойствам цитохрома с. Цитохром с исследовался очень детально, но его динамические свойства нельзя считать полностью установленными и объясненными.
Строение и функциональная организация митохондрий явились предметом интенсивного изучения. Тем не менее, многие важнейшие вопросы, сюда относящиеся, пока не имеют ответа. Специфическая мембранная структура мигохондрий, присутствие в них автономной программы синтеза белков (ДНК), механохи - мическая активность митохондрий непосредственно связаны с их ролью «силовых станций» клетки. Дальнейшее развитие физики митохондрий требует целостного подхода.
Общая теория электронного транспорта в локализованной системе биологического окисления интенсивно развивается. Предложены содержательные кинетические модели, начаты теоретические исследования, основанные на рассмотрении элект - ронно-конформационных взаимодействий. Эти проблемы представляют первостепенный интерес для биофизики.
Необходимо установить молекулярную природу биологического окисления, осуществляемого в результате сопряжения ферментативных, транспортных и механохимических процессов. Митохондрия есть место интегрирования широкой совокупности мо - лекулярно-биологических явлений, целостная система, требующая всестороннего изучения - теоретической и экспериментальной разборки и сборки «черного ящика».
Биологическое окисление , происходящее в живом организме, по сути, является процессом, обратным фотосинтезу. В ходе реакций биологического окисления высокоэнергетические электроны, находящиеся в молекулах углеводов и других биологических соединений, скатываются на уровень с наименьшей энергией, когда связываются с кислородом в молекуле воды. Энергия, отдаваемая ими при этом, используется для образования макроэргических фосфатных связей. Поток электронов, движущихся по ступеням процесса биологического окисления, - это не что иное, как слабый электрический ток.
Биологическое окисление это процесс окисления биологических веществ с выделением энергии.
Тканевое дыхание – процесс поглощения кислорода (О 2) тканями при окислении органического субстрата с выделением углекислого газа (СО 2) и воды (Н 2 О).
Главными источниками СО 2 является реакции декарбоксилирования пировиноградной и альфа-кетоглутаровой кислот. Еще один источник – это процесс декарбоксилирование аминокислот, который катализируется пиридоксаль-зависимыми ферментами.
Окислительное фосфорилирование это синтез АТФ сопряженный с тканевым дыханием.
Основным топливом при биологическом окислении является водород. Известно, что реакция окисление водорода кислородом в газовой среде сопровождается выделением большого количества энергии, сопровождаемым взрывом и пламенем. Эволюция живых организмов привела к тому, что реакция окисления водорода до воды оказалась разделенной на отдельные этапы, что обеспечивает постепенное высвобождение энергии в процессе биологического окисления. При этом часть полученной энергии рассеивается в виде тепла (около 60%) а другая часть (около 40%) аккумулируется в молекулах АТФ.
Макроэргические соединения – это вещества, содержащие богатые энергией связи.
Макроэргическая связь обозначается символом ∼ (знак «тильда»). Понятие макроэргическая связь довольно условно и применяется для обозначения тех связей, которые гидролизуются в водной среде с выделением значительной энергии. Так, гидролиз концевой фосфоангидридной связи АТФ (АТФ + Н 2 О → АДФ + Фн), ведет к освобождению 34,5 кДж/моль энергии).
Однако если реакция протекает в неводной среде (например, в липидном слое мембран), то образование и разрушение АТФ протекает без больших затрат энергии.
Вещества гидролиз которых, приводит к высвобождению более 21 кДж/моль энергии относят к высокоэнергетическим (макроэргам ), а вещества освобождающие меньшие количества энергии - к низкоэнергетическим. К макроэргам относят: АТФ, другие трифосфаты нуклеозидов (ГТФ, ЦТФ, УТФ, ТТФ), аргининфосфат, креатинфосфат, ацетилфосфат, 1,3дифосфоглицерат, фосфоенолпируват и др. К низкоэнергетическим - глюкозо-6-фосфат, глюкозо-1-фосфат, глицерофосфат и другие. Уникальная роль АТФ состоит в том, что она имеет промежуточное значение энергии гидролиза и выполняет роль связующего звена (разменной монеты) между высоко- и низкоэнергетическими соединениями.
Слайд 2
Совокупность окислительных реакций, происходящих в биологических объектах и обеспечивающих их энергией и метаболитами для осуществления процессов жизнедеятельности, называется биологическим окислением.
Слайд 3
Функции биологического окисления
Энергетическое обеспечение:
- поддержания температуры тела;
- биолюминесценции (свечения);
- химических синтезов;
- осмотических явлений;
- электрических процессов;
- механической работы.
- Синтез важнейших (ключевых) метаболитов.
- Регуляция обмена веществ.
- Устранение вредных для клетки продуктов обмена (шлаков).
- Детоксикация проникших в организм чуждых соединений – ксенобиотиков (пестицидов, препаратов бытовой химии, лекарственных средств, промышленных загрязнений и т.п.).
Слайд 4
Ферменты биологического окисления
- Разнообразные реакции биологического окисления ускоряются многочисленными ферментами оксидоредуктазами, которые, как правило, встроены в биологические мембраны, причем очень часто в виде ансамблей.
Их разделяют на 5 групп:
- Оксидазы (катализируют удаление водорода из субстрата, используя при этом в качестве акцептора водорода только кислород)
Слайд 5
Аэробные дегидрогеназы (в отличие от оксидаз они могут использовать в качестве акцептора водорода не только кислород, но и искусственные акцепторы).
Слайд 6
- Анаэробные дегидрогеназы (не способны использовать кислород в качестве акцептора водорода)
- Выполняют две главные функции:
- Перенос водорода с одного субстрата на другой
- Компонент дыхательной цепи, обеспечивающий транспорт электронов от субстрата на кислород
Слайд 7
- Гидроксипероксидазы (используют в качестве субстрата перекись водорода или органические перекиси)
- Оксигеназы (катализируют прямое введение кислорода в молекулу субстрата)
Слайд 8
Типы биологического окисления
- Существуют 2 типа биологического окисления:
- Свободное окисление
- Окисление, не сопряженное с фосфорилированием АДФ и не сопровождающееся трансформацией энергии, выделяющейся при окислении, в энергию макроэргических связей.
- При свободном окислении высвобождающаяся энергия переходит в тепловую и рассеивается.
- Этот тип биологического окисления осуществляется двумя путями:
- субстратное фосфорилирование
- окислительное фосфорилирование
Слайд 9
Свободное окисление
Слайд 10
- Реакции свободного окисления органических соединений в живой природе и ускоряющие их ферментные системы многообразны. Этим путем непосредственно окисляются не только многочисленные природные и неприродные субстраты, но и восстановленные коферменты (НАДН, НАДФН, ФАД·Н2 и др.), образовавшиеся при действии первичных и вторичных дегидрогеназ.
- Реакции свободного окисления протекают в цитозоле, на мембранах различных субклеточных структур, в ядерном аппарате клетки. Основным средоточием их являются мембраны эндоплазматической сети (ЭПС).
- Так как мембраны ЭПС при гомогенизации клеток и фракционировании субклеточных частиц гомогената дают фракцию микросом, то реакции окисления на мембранах ЭПС называются микросомальным окислением.
Слайд 11
Особенности микросомальной дыхательной цепи
- Несмотря на наличие ферментов цепи переноса электронов, ни в одном пункте этой цепи не происходит сопряжения с фосфорилированием АДФ.
- Своеобразие структуры и функциональной активности цитохромовb5 и Р–450, входящих в ее состав.
- Высокое сродство терминальной оксидазы микросомальных цепей к кислороду, позволяющее ей конкурировать за кислород с митохондриальнойцитохромоксидазой.
Слайд 12
Ферменты свободного окисления
Слайд 13
- Пирокатехаза (катехол: кислород -1,2-оксидоредуктаза дециклизующая)
- Она содержит в активном центре два прочно связанных атома Fе, которые, согласно О. Хайаиши, соединяются с молекулярным кислородом в комплекс, где кислород далее активируется:
- Fe2+ + О2 → Fе2+О2 → Fе3+О2–
Слайд 14
Слайд 15
Примеры диоксигеназных реакций
Слайд 16
Свободное окисление при участии монооксигеназ
Слайд 17
Окисление, сопряженное с фосфорилированием АДФ
Слайд 18
Слайд 19
Субстратное фосфорилирование
Слайд 20
Субстратное фосфорилирование – такой вид биологического окисления, при котором: макроэргическая связь возникает в момент непосредственного окисления субстрата, затем тем или иным путем передается на фосфатный остаток, который, в свою очередь, используется для фосфорилирования АДФ, т.е. синтеза АТФ. Окисление, сопряженное с фосфорилированием АДФ на уровне субстрата.
Слайд 21
Примеры реакций субстратного фосфорилирования
- При окислении 3-фосфоглицеринового альдегида (3-ФГА) в 2-фосфоглицериновую кислоту (2-ФГК) – гликолиз;
- При превращении фосфоенолпировиноградной кислоты (ФЕП) в пировиноградную (пируват, ПВК) – гликолиз;
- При превращении -кетоглутаровой кислоты в янтарную (реакция цикла Кребса).
Слайд 22
Окислительное декарбоксилирование кетоглутаровой кислоты
На этом участке цикла Кребса донором электронов является оксиацетилтиаминпирофосфат; акцептором электронов – липоевая кислота.
Слайд 23
Слайд 24
Слайд 25
На следующих этапах в реакцию вступает фермент
Слайд 26
Окислительное фосфорилирование
Слайд 27
- Это сопряжение окисления с синтезом АТФ, когда атомы водорода с коферментов дегидрогеназ, принимающих участие в окислении субстратов, передаются в оксидоредуктазную цепь, где сопряжено с переносом ионов Н+ и электронов на молекулярный кислород происходит активирование неорганического фосфата и при его посредстве – фосфорилирование АДФ с образованием АТФ
- Окисляемый субстрат в этом случае непосредственного участия в активировании неорганического фосфата не принимает
- Сопряжение окисления с фосфорилированием идет главным образом на внутренних мембранах митохондрий
Слайд 28
Окислительно-восстановительные ферменты
Слайд 29
Пиридиновые дегидрогеназы
- Коферменты – НАД и НАДФ
- Универсальный донор атомов Н для дыхательной цепи ферментов – НАДН2
- Если при окислении субстрата возникает НАДФН2, то осуществляется реакция:
- НАДФН2 + НАД ⇄ НАДФ + НАДН2
Слайд 30
Особенности реакций с участием пиридиновых дегидрогеназ
- Легкая обратимость.
- Коферменты легко отделяются от белковой части, обладают высокой подвижностью, что позволяет им переносить атомы Н, ионы Н+ и электроны из одной части клетки в другую.
- НАД и НАДФ способны принимать атомы Н от большого числа субстратов, окислительно-восстановительные потенциалы которых ниже (-0,32В).
Слайд 31
Флавиновые дегидрогеназы
- Коферменты – ФМН и ФАД.
- Флавиновые ферменты являются акцепторами атомов водорода и осуществляют перенос их от НАДН2:
- НАДН2 + ФАД ⇄ НАД + ФАДН2.
- В некоторых случаях (при окислении янтарной кислоты в цикле Кребса или при окислении жирных кислот) флавиновые ферменты могут играть роль первичных дегидрогеназ.
- ФМН и ФАД очень прочно связаны с апоферментом и не отщепляются от него ни на одной стадии каталитического цикла.
- Активной частью молекул ФАД и ФМН является изоаллоксазиновое кольцо рибофлавина, к атомам азота которого могут присоединяться 2 атома водорода:
Слайд 36
- Самой примечательной особенностью дыхательной цепи ферментов является наличие в ней участков, где соседние компоненты резко отличаются значениями окислительно-восстановительных потенциалов.
- Именно здесь происходит сопряжение окисления с фосфорилированием АДФ, т.к. разность энергетических уровней электрона, транспортируемого с огромной скоростью, вполне достаточна для синтеза макроэргической связи и составляет 51 кДж для I, 36 кДж – для IIи 80,7 кДж – для III точки сопряжения.
Слайд 37
Хемиосмотическая гипотеза
- Питер Митчелл, Владимир Петрович Скулачев
- Реакции, сопровождающиеся расходованием или образованием Н+, протекают на внутренней мембране митохондрий таким образом, что протоны переносятся с внутренней мембраны на внешнюю, т.е. перенос электронов сопровождается возникновением трансмембранного градиента концентрации ионов Н+ – совершением осмотической работы.
- Этот градиент, создающий разность химических () и электрических () потенциалов, является источником энергии для протекания эндэргонического процесса образования АТФ.
- АТФаза является ферментом, способным использовать градиент концентрации ионов Н+ для обращения процесса гидролиза АТФ.
Слайд 38
Посмотреть все слайды